
Журнал «Здоровье ребенка» Том 14, №8, 2019
Вернуться к номеру
Медикаментозное влияние на диспергирование биопленки. Аминокислоты и их дериваты
Авторы: Абатуров А.Е.(1), Крючко Т.А.(2)
(1) — ГУ «Днепропетровская медицинская академия МЗ Украины», г. Днепр, Украина
(2) — ВГУЗУ «Украинская медицинская стоматологическая академия», г. Полтава, Украина
Рубрики: Педиатрия/Неонатология
Разделы: Справочник специалиста
Версия для печати
У науковому огляді подана значущість деяких амінокислот у процесі диспергування бактеріальних біоплівок. Для написання статті виконувався пошук інформації з використанням баз даних Scopus, Web of Science, MedLine, PubMed, Google Scholar, EMBASE, Global Health, The Cochrane Library, CyberLeninka. Відображено вплив амінокислот як нутрієнтів на пізні стадії життя біоплівки, представлено вплив D-енантіомерів амінокислот, триптофану та його дериватів на диспергування біоплівки. Підкреслено, що D-амінокислоти, а саме D-тирозин, D-метіонін, D-лейцин, D-триптофан, перешкоджають адгезії амілоїдних волокон, які утримують в агрегації бактерії біоплівок. D-амінокислоти інгібують експресію генів, що беруть участь у продукції компонентів матриксу біоплівки, і таким чином реалізують свою антибіоплівкову дію. Розкрито здатність триптофану попереджати розвиток і викликати диспергування біоплівок, зокрема асоційованих з бактеріями Pseudomonas aeruginosa. Зазначено, що роль антранілатів полягає в порушенні структури біоплівки і стимулюванні відриву ділянок від біоплівок, сформованих бактеріями Pseudomonas aeruginosa. Антранілати можуть сприяти перериванню хронізації інфекційного процесу. Припущено, що використання амінокислот і їх похідних, створення нових хімічних сполук, які імітують структури молекул амінокислот або змінюють активність синтезу і деградації певних амінокислот, дозволить цілеспрямовано управляти циклом життя біоплівок деяких видів патогенних бактерій, що буде сприяти виліковуванню інфекційних рецидивуючих і хронічних захворювань. Подано перспективу розробки нових препаратів на основі антранілової кислоти і галогенованих індолів, що сприяють бактеріальній ерадикації при інфекціях респіраторного тракту, асоційованих з розвитком біоплівок бактерій антибіотикорезистентних бактеріальних агентів.
В научном обзоре представлено значение некоторых аминокислот в процессе диспергирования бактериальных биопленок. Для написания статьи осуществлялся поиск информации с использованием баз данных Scopus, Web of Science, MedLine, PubMed, Google Scholar, EMBASE, Global Health, The Cochrane Library, CyberLeninka. Отражено влияние аминокислот как нутриентов на поздние стадии жизни биопленки, представлено влияние D-энантиомеров аминокислот, триптофана и его дериватов на диспергирование биопленки. Подчеркнуто, что D-аминокислоты, а именно D-тирозин, D-метионин, D-лейцин, D-триптофан, препятствуют адгезии амилоидных волокон, которые удерживают в агрегации бактерии биопленок. D-аминокислоты ингибируют экспрессию генов, участвующих в продукции компонентов матрикса биопленки, и таким образом реализуют свое антибиопленочное действие. Раскрыта способность триптофана предупреждать развитие и вызывать диспергирование биопленок, в частности, ассоциированных с бактериями Pseudomonas aeruginosa. Указано, что роль антранилатов заключается в нарушении структуры биопленки и стимулировании отрыва участков от биопленок, сформированных бактериями Pseudomonas aeruginosa. Антранилаты могут способствовать прерыванию хронизации инфекционного процесса. Предположено, что использование аминокислот и их дериватов, создание новых химических соединений, которые имитируют структуры молекул аминокислот или изменяют активность синтеза и деградации определенных аминокислот, позволит целенаправленно управлять циклом жизни биопленок некоторых видов патогенных бактерий, что будет способствовать излечению инфекционных рецидивирующих и хронических заболеваний. Представлена перспектива разработки новых препаратов на основе антраниловой кислоты и галогенерированных индолов, способствующих бактериальной эрадикации при инфекциях респираторного тракта, ассоциированных с развитием биопленок бактерий антибиотикорезистентных бактериальных агентов.
The scientific review presents the significance of some amino acids in the process of dispersing of bacterial biofilms. To write the article, information was searched using Scopus, Web of Science, MedLine, PubMed, Google Scholar, EMBASE, Global Health, the Cochrane Library, CyberLeninka. The influence of amino acids as nutrients on the late stages of biofilm life is considered, the effect of the D-enantiomers of amino acids on tryptophan and its derivatives on the biofilm dispersion is presented. It is emphasized that D-amino acids, namely, D-tyrosine, D-methionine, D-leucine, D-tryptophan, prevent the adhesion of amyloid fibers, which keep the aggregation of the biofilms. D-amino acids inhibit the expression of genes involved in the production of components of the biofilm matrix, and thus realize their antibiotic effect. The ability of tryptophan to prevent the development and dispersion of biofilms, in particular those associated with the bacteria Pseudomonas aeruginosa, is highlighted. It is indicated that the role of anthranilates is to disrupt the structure of the biofilm and to stimulate the separation of sites from biofilms formed by the bacteria Pseudomonas aeruginosa. Anthranilates can help interrupt the chronicity of the infectious process. It is assumed that the use of amino acids and their derivatives, the creation of new chemical compounds that mimic the structure of amino acid molecules or alter the activity of the synthesis and degradation of certain amino acids will allow purposefully to control the life cycle of biofilms of certain types of pathogenic bacteria, which will contribute to the recovery of infectious recurrent and chronic diseases. The paper presents the prospect of developing new drugs based on anthranilic acid and halogenated indoles that promote bacterial eradication in respiratory tract infections associated with the development of bacterial biofilms of antibiotic-resistant bacterial agents.
диспергування біоплівки; респіраторний тракт; амінокислоти та їх деривати; рецидивуючі та хронічні інфекційно-запальні захворювання; огляд
диспергирование биопленки; респираторный тракт; аминокислоты и их дериваты; рецидивирующие и хронические инфекционно-воспалительные заболевания; обзор
biofilm dispersion; respiratory tract; amino acids and their derivatives; recurrent and chronic infectious and inflammatory diseases; review
Введение
Аминокислоты и их дериваты активно влияют на процесс диспергирования биопленки. Концентрация аминокислот как ключевых бактериальных нутриентов определяет переход от одной формы жизни бактериальной колонии к другой. Изменение доступности питательных веществ способствует переходу от оседлой формы жизни бактерий к планктонной или наоборот [32]. Также стресс-ассоциированная продукция D-аминокислот способствует диспергированию биопленок [7, 10]. Из совокупности аминокислот особое влияние на диспергирование биопленок оказывают дериваты триптофана — антранилаты [17] и индол [16]. Медикаментозное управление обеспеченностью питательными веществами, синтезом D-аминокислот и продукцией дериватов триптофана может являться новым стратегическим направлением лечения бактериальных инфекций, которые сопровождаются развитием биопленки [26].
Нутриенты (глутамат и глутамин)
Известно, что формирование бактериальных биопленок зависит от обеспечения глутамином (глутаминовая кислота) или глутаматом в качестве источника азота. Глутамин и глутамат поступают в бактерии из окружающей среды или синтезируются непосредственно ферментными механизмами бактерий. Глутамин образуется при помощи глутаминсинтетазы из глутамата и аммиака, а глутамат — при катализе глутамина. Представляет интерес то, что жизнедеятельность бактерий различных участков биопленки характеризуется метаболической зависимостью. Так, бактерии, расположенные в периферических участках биопленки, получают глутамат из окружающей среды, но не способны синтезировать аммиак. Аммиак синтезируют исключительно бактерии, локализованные в центральных участках биопленки. Достаточная обеспеченность аммиаком способствует конверсии глутамата в глутамин, который является преимущественным нутриентом для бактерий. Таким образом, рост периферически расположенных бактерий рестриктирован уровнем активности продукции аммиака бактериями центральных участков биопленки, дефицит аммиака приводит к ограничению потребления глутамата из окружающей среды [27].
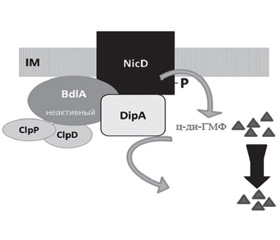
Ankita Basu Roy и Karin Sauer [5] сформулировали положение, согласно которому диспергирование бактерий из биопленок активируется в ответ на повышение уровня питательных веществ в окружающей среде. Авторы продемонстрировали, что нутриент-индуцированная циклаза внутренней мембраны D (nutrient-induced cyclase D — NicD) представляет собой дигуанилатциклазу (diguanylate cyclases — DGC), принадлежащую к семейству семи трансмембранных (7TM) рецепторов и участвующую в синтезе циклического дигуанозинмонофосфата (cyclic diguanyl monophosphate — c-di-GMP/ц-ди-ГМФ). Дигуанилатциклаза NicD, регулируя активность синтеза ц-ди-ГМФ, контролирует процесс диспергирования биопленки. При увеличении концентраций глутамата протеин NicD дефосфорилируется, что сопровождается увеличением уровня его каталитической активности. Активированная дигуанилатциклаза NicD способствует протеолизу и активации протеина BdlA и транзиторному повышению уровня концентрации ц-ди-ГМФ. В свою очередь, фосфорилированный BdlA активирует фосфодиэстеразу DipA, что приводит к уменьшению концентрации ц-ди-ГМФ и к диспергированию биопленки (рис. 1).
/107-1.jpg)
Tal Hassanov и соавт. [12] показали, что манипуляция активностью метаболических путей глутамин/глутамат обладает терапевтическим потенциалом, который может быть использован при лечении инфекционных заболеваний, ассоциированных с формированием биопленки. Так, авторы продемонстрировали, что два ингибитора синтеза глутамина и глутамата (6-диазо-5-оксо-L-норлейцин) и ингибитор глутаматной оксалоацетатной трансаминазы и аспартатаминотрансферазы (аналог глутамина и аминооксиуксусной кислоты) препятствуют развитию в основном периферических участков биопленки.
D-аминокислоты
Молекулы аминокислот (кроме глицина) могут пребывать в лево- (L-аминокислоты) и правовращающейся форме (D-аминокислоты). Большинство энантиомеров натуральных аминокислот бактерий находятся в L-форме и являются компонентами протеинов, а небольшая часть аминокислот, представленная D-формами, участвует в формировании непротеиновых компонентов бактериальных стенок. D-аминокислоты высвобождаются при истощении питательных веществ и индуцируют диспергирование сформированных бактериальных биопленок [2, 7].
Продемонстрировано, что D-аминокислоты, в частности D-тирозин, D-метионин, D-лейцин, D-триптофан, взаимодействуя с пептидогликаном, препятствуют адгезии амилоидных волокон, которые удерживают в агрегации бактерии биопленок [19, 28]. Также D-аминокислоты реализуют свое антибиопленочное действие за счет ингибирования экспрессии генов, участвующих в продукции компонентов матрикса биопленки [24, 31]. D-аминокислоты могут влиять на диспергирование биопленок, сформированных бактериями Staphylococcus aureus и Pseudomonas aeruginosa [1, 2, 13].
Andrew J. Harmata и соавт. [11] полагают, что локальная доставка D-аминокислот будет эффективной терапевтической стратегией при лечении инфекционных заболеваний, ассоциированных с развитием биопленки.
Carlos J. Sanchez и соавт. [30], оценивая in vitro диспергирующее влияние D-аминокислот на биопленки, сформированные клиническими изолятами из раневых поверхностей бактерий Staphylococcus aureus и Pseudomonas aeruginosa, определили, что D-метионин, D-триптофан при концентрациях ≥ 5 ммоль индуцируют распад биопленки. Комбинации D-аминокислот и антибиотиков (клиндамицина, цефазолина, оксациллина, рифампина, ванкомицина для бактерий Staphylococcus aureus и амикацина, колистина, ципрофлоксацина, имипенема, цефтазидима для бактерий Pseudomonas aeruginosa) характеризуются высокой антибиопленочной и эрадикационной активностью.
Триптофан и его дериваты
Триптофан является мощным агентом, предупреждающим развитие и вызывающим диспергирование биопленок, в частности, ассоциированных с бактериями Pseudomonas aeruginosa [6]. Установлено, что триптофан подавляет экспрессию кворум-сенсинг-ассоциированных генов lasR, lasB и lasI бактерий Pseudomonas aeruginosa [8]. Kenneth S. Brandenburg и соавт. [6] продемонстрировали, что D- и L-изоформы триптофана обладают практически одинаковой эффективностью при ингибировании образования биопленки бактериями Pseudomonas aeruginosa, а смесь D- и L-изоформ триптофана превосходит по силе антибиопленочного действия и D-, и L-энантиомеры триптофана. Авторы предполагают, что триптофан ингибирует образование и усиливает диспергирование биопленки за счет усиления подвижности бактерий Pseudomonas aeruginosa. Учитывая, что биопленки содержат многочисленный пул немобильных бактерий, тириптофан-индуцированное усиление подвижности бактериальных единиц может способствовать отделению бактерий или бактериального агрегата от биопленки. Установлено, что триптофан при концентрациях, ингибирующих образование биопленки (от 1 до 10 ммоль), не является цитотоксичным веществом для клеток человека.
Дериваты триптофана (антраниловая кислота, индол) также индуцируют диспергирование биопленок [26].
Антраниловая кислота
Некоторые бактерии, в частности бактерии Pseudomonas aeruginosa, метаболизируют триптофан при помощи ферментов триптофан-2,3-диоксигеназы (ген kynA), кинуренинформамидазы (ген kynB) и кинурениназы (ген kynU) в антраниловую кислоту, которая необходима для формирования биопленки [20]. Мутантные бактерии Pseudomonas aeruginosa, не способные деградировать антранилаты, образуют значительно менее массивные биопленки, чем дикие бактериальные формы [9]. Антранилаты нарушают структуру биопленки и стимулируют отрыв участков от биопленок, сформированных бактериями Pseudomonas aeruginosa, за счет снижения уровня внутриклеточной концентрации ц-ди-ГМФ и подавления активности экспрессии генов, участвующих в продукции компонентов экстрацеллюлярного матрикса [18]. Согласно результатам исследования, проведенного Xi-Hui Li и соавт. [25], антранилаты оказывают ингибирующее действие на развитие биопленок, организованных такими микроорганизмами, как бактерии Pseudomonas aeruginosa, Bacillus subtilis, Vibrio vulnificus, Salmonella enterica. Авторы показали, что, во-первых, эффективность ингибирования биопленки у антранилатов достоверно выше, чем у L-триптофана, и, во-вторых, антранилаты не оказывают цитотоксического эффекта на клетки макроорганизма.
Soo-Kyoung Kim и соавт. [18] показали, что антранилаты оказывают влияние на экспрессию гена Psl бактерий Pseudomonas aeruginosa, продукт которого играет ключевую роль в первоначальной адгезии бактерий к абиотическим и биотическим поверхностям, и на синтез альгинатов, принимающих участие в поздних стадиях формирования биопленки. На ранней стадии инфекционного процесса антранилаты усиливают экспрессию гена Psl бактерий Pseudomonas aeruginosa, способствуя первичному прикреплению микроорганизмов, а в позднем периоде формирования биопленки антранилаты ингибируют как экспрессию гена Psl, так и продукцию альгината. Авторы подчеркивают, что действие антранилата на транскрипцию оперонов Psl и альгината противоположно фактору транскрипции AmrZ, который, как известно, подавляет активность генов, ответственных за подвижность бактерий Pseudomonas aeruginosa [3]. Фактор транскрипции AmrZ индуцирует продукцию альгината, но репрессирует транскрипцию гена Psl, что способствует переходу бактерий Pseudomonas aeruginosa от колонизации и ранней стадии биопленки к формированию зрелой биопленки, характерной для хронического течения инфекционных заболеваний [15]. Таким образом, антранилаты могут выступать в качестве саногенического фактора, прерывающего хронизацию инфекционного процесса.
Учитывая, что антраниловая кислота является естественным соединением с простой молекулярной структурой, синтез которой не требует значительных финансовых затрат, полагают, что она представляет собой многообещающее соединение, на основе которого будут созданы лекарственные средства, способствующие бактериальной эрадикации при инфекциях респираторного тракта, ассоциированных с развитием биопленок бактерий Pseudomonas aeruginosa.
Индол и производные
Индол является метаболитом триптофана и продуцируется многочисленными (около 85) видами бактерий, геном которых содержит ген TnaA, кодирующий триптофаназу. Доказано, что эукариотические клетки не продуцируют индол [22, 23]. Однако индол, вырабатываемый бактериями, может оказывать клинически значимое влияние на состояние здоровья человека. В частности, показано, что у людей, у которых микробиота кишечника продуцирует избыточное количество индола, отмечается высокая вероятность развития тревожности и расстройств настроения [14].
Наличие или отсутствие гена триптофаназы определяет эффекты действия индола на биопленку бактерий. Так, показано, что индол снижает образование биопленки, организованной индол-продуцирующими бактериями Escherichia coli, и в то же время он усиливает развитие биопленки индол-непродуцирующими бактериями Pseudomonas aeruginosa [18].
Установлено, что индол способствует диспергированию биопленки, сформированной бактериями Escherichia coli, за счет повышения подвижности [29] и подавления активности адгезии микроорганизмов к эпителиальным клеткам [4].
Jin-Hyung Lee и соавт. [21] провели исследования способности ингибировать образование персистентных форм и биопленок грамотрицательных бактерий Escherichia coli и грамположительных бактерий Staphylococcus aureus у 36 различных производных индола. Авторами установлено, что галогенированные индолы 5-йодиндол, 4-фториндол, 7-хлориндол и 7-броминдол обладают выраженной антибиопленочной активностью. В отличие от других галогенированных индолов 5-йодиндол не индуцирует образование персистирующих бактерий, а ингибирует продукцию такого фактора вирулентности бактерий Staphylococcus aureus, как иммуноинвазивный каротиноидный стафилоксантин. Полагают, что галогенированные индолы обладают терапевтическим потенциалом, который может быть использован при эрадикации антибиотикорезистентных бактериальных агентов.
Заключение
Невозможно переоценить вклад аминокислот в любую сферу жизнедеятельности бактерий, в том числе и в процесс диспергирования биопленок. Изменение активности нутриент-ассоциированных сигнальных путей, концентрации D-форм некоторых аминокислот, триптофана и его дериватов оказывает влияние на формирование патогенных бактериальных биопленок. Использование аминокислот и их дериватов, создание новых химических соединений, которые имитируют структуры молекул аминокислот или изменяют активность синтеза и деградации определенных аминокислот, позволит целенаправленно управлять циклом жизни биопленок некоторых видов патогенных бактерий, что будет способствовать излечению инфекционных рецидивирующих и хронических заболеваний.
Конфликт интересов. Авторы заявляют об отсутствии какого-либо конфликта интересов и собственной финансовой заинтересованности при подготовке данной статьи.
1. Aliashkevich A., Alvarez L., Cava F. New Insights Into the Mechanisms and Biological Roles of D-Amino Acids in Complex Eco-Systems. Front. Microbiol. 2018 Apr 6. 9. 683. doi: 10.3389/fmicb.2018.00683.
2. Ampornaramveth R.S., Akeatichod N., Lertnukkhid J., Songsang N. Application of D-Amino Acids as Biofilm Dispersing Agent in Dental Unit Waterlines. Int. J. Dent. 2018 Jan 14. 2018. 9413925. doi: 10.1155/2018/9413925.
3. Baltrus D.A., Dougherty K., Diaz B., Murillo R. Evolutionary Plasticity of AmrZ Regulation in Pseudomonas. mSphere. 2018 Apr 18. 3(2). pii: e00132-18. doi: 10.1128/mSphere.00132-18.
4. Bansal T., Englert D., Lee J. et al. Differential effects of epinephrine, norepinephrine, and indole on Escherichia coli O157:H7 chemotaxis, colonization, and gene expression. Infect. Immun. 2007 Sep. 75(9). 4597-607. doi: 10.1128/IAI.00630-07.
5. Basu Roy A., Sauer K. Diguanylate cyclase NicD-based signalling mechanism of nutrient-induced dispersion by Pseudomonas aeruginosa. Mol. Microbiol. 2014 Nov. 94(4). 771-93. doi: 10.1111/mmi.12802.
6. Brandenburg K.S., Rodriguez K.J., McAnulty J.F. et al. Tryptophan inhibits biofilm formation by Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2013 Apr. 57(4). 1921-5. doi: 10.1128/AAC.00007-13.
7. Cava F., Lam H., de Pedro M.A., Waldor M.K. Emerging knowledge of regulatory roles of D-amino acids in bacteria. Cell. Mol. Life Sci. 2011 Mar. 68(5). 817-31. doi: 10.1007/s00018-010-0571-8.
8. Chakraborty P., Daware A.V., Kumari M. et al. Free tryptophan residues inhibit quorum sensing of Pseudomonas aeruginosa: a potential approach to inhibit the development of microbial biofilm. Arch. Microbiol. 2018 Jul 23. doi: 10.1007/s00203-018-1557-4.
9. Costaglioli P., Barthe C., Claverol S. et al. Evidence for the involvement of the anthranilate degradation pathway in Pseudomonas aeruginosa biofilm formation. Microbiologyopen. 2012 Sep. 1(3). 326-39. doi: 10.1002/mbo3.33.
10. Fleming D., Rumbaugh K.P. Approaches to Dispersing Medical Biofilms. Microorganisms. 2017 Apr 1. 5(2). pii: E15. doi: 10.3390/microorganisms5020015.
11. Harmata A.J., Ma Y., Sanchez C.J. et al. D-amino acid inhibits biofilm but not new bone formation in an ovine model. Clin. Orthop. Relat. Res. 2015 Dec. 473(12). 3951-61. doi: 10.1007/s11999-015-4465-9.
12. Hassanov T., Karunker I., Steinberg N. et al. Novel antibiofilm chemotherapies target nitrogen from glutamate and glutamine. Sci Rep. 2018 May 8. 8(1). 7097. doi: 10.1038/s41598-018-25401-z.
13. Hochbaum A.I., Kolodkin-Gal I., Foulston L. et al. Inhibitory effects of D-amino acids on Staphylococcus aureus biofilm development. J. Bacteriol. 2011 Oct. 193(20). 5616-22. doi: 10.1128/JB.05534-11.
14. Jaglin M., Rhimi M., Philippe C. et al. Indole, a Signaling Molecule Produced by the Gut Microbiota, Negatively Impacts Emotional Behaviors in Rats. Front Neurosci. 2018 Apr 9. 12. 216. doi: 10.3389/fnins.2018.00216.
15. Jones C.J., Ryder C.R., Mann E.E., Wozniak D.J. AmrZ modulates Pseudomonas aeruginosa biofilm architecture by directly repressing transcription of the psl operon. J. Bacteriol. 2013 Apr. 195(8). 1637-44. doi: 10.1128/JB.02190-12.
16. Kim J., Park W. Indole: a signaling molecule or a mere metabolic byproduct that alters bacterial physiology at a high concentration? J. Microbiol. 2015 Jul. 53(7). 421-8. doi: 10.1007/s12275-015-5273-3.
17. Kim S.K., Lee J.H. Biofilm dispersion in Pseudomonas aeruginosa. J. Microbiol. 2016 Feb. 54(2). 71-85. doi: 10.1007/s12275-016-5528-7.
18. Kim S.K., Park H.Y., Lee J.H. Anthranilate deteriorates the structure of Pseudomonas aeruginosa biofilms and antagonizes the biofilm-enhancing indole effect. Appl. Environ. Microbiol. 2015 Apr. 81(7). 2328-38. doi: 10.1128/AEM.03551-14.
19. Kolodkin-Gal I., Romero S. Cao et al. D-amino acids trigger biofilm disassembly. Science. 2010 Apr 30. 328(5978). 627-9. doi: 10.1126/science.1188628.
20. Kurnasov O., Jablonski L., Polanuyer B. et al. Aerobic tryptophan degradation pathway in bacteria: novel kynurenine formamidase. FEMS Microbiol Lett. 2003 Oct 24. 227(2). 219-27.
21. Lee J.H., Kim Y.G., Gwon G. et al. Halogenated indoles eradicate bacterial persister cells and biofilms. AMB Express. 2016 Dec. 6(1). 123. doi: 10.1186/s13568-016-0297-6.
22. Lee J.H., Lee J. Indole as an intercellular signal in microbial communities. FEMS Microbiol Rev. 2010 Jul. 34(4). 426-44. doi: 10.1111/j.1574-6976.2009.00204.x.
23. Lee J.-H., Wood T.K., Lee J. Roles of indole as an interspecies and interkingdom signaling molecule. Trends in microbiology. 2015. 23. 707-718. doi: 10.1016/j.tim.2015.08.001.
24. Leiman S.A., May J.M., Lebar M.D. et al. D-amino acids indirectly inhibit biofilm formation in Bacillus subtilis by interfering with protein synthesis. J. Bacteriol. 2013 Dec. 195(23). 5391-5. doi: 10.1128/JB.00975-13.
25. Li X.H., Kim S.K., Lee J.H. Anti-biofilm effects of anthranilate on a broad range of bacteria. Sci Rep. 2017 Aug 17. 7(1). 8604. doi: 10.1038/s41598-017-06540-1.
26. Li X.H., Lee J.H. Antibiofilm agents: A new perspective for antimicrobial strategy. J. Microbiol. 2017 Oct. 55(10). 753-766. doi: 10.1007/s12275-017-7274-x.
27. Liu J., Prindle A., Humphries J. et al. Metabolic co-dependence gives rise to collective oscillations within biofilms. Nature. 2015 Jul 30. 523(7562). 550-4. doi: 10.1038/nature14660.
28. Oppenheimer-Shaanan Y., Steinberg N., Kolodkin-Gal I. Small molecules are natural triggers for the disassembly of biofilms. Trends Microbiol. 2013 Nov. 21(11). 594-601. doi: 10.1016/j.tim.2013.08.005.
29. Pringle S.L., Palmer K.L., McLean R.J. Indole production provides limited benefit to Escherichia coli during co-culture with Enterococcus faecalis. Arch. Microbiol. 2017 Jan. 199(1). 145-153. doi: 10.1007/s00203-016-1289-2.
30. Sanchez C.J. Jr, Akers K.S., Romano D.R. et al. D-amino acids enhance the activity of antimicrobials against biofilms of clinical wound isolates of Staphylococcus aureus and Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2014 Aug. 58(8). 4353-61. doi: 10.1128/AAC.02468-14.
31. Sanchez Z., Tani A., Kimbara K. Extensive reduction of cell viability and enhanced matrix production in Pseudomonas aeruginosa PAO1 flow biofilms treated with a D-amino acid mixture. Appl. Environ. Microbiol. 2013 Feb. 79(4). 1396-9. doi: 10.1128/AEM.02911-12.
32. Sauer K., Cullen M.C., Rickard A.H. et al Characterization of nutrient-induced dispersion in Pseudomonas aeruginosa PAO1 biofilm. J. Bacteriol. 2004 Nov. 186(21). 7312-26. doi: 10.1128/JB.186.21.7312-7326.2004.