
Журнал «Здоровье ребенка» 3 (63) 2015
Вернуться к номеру
Значення однонуклеотидних поліморфізмів в генах mTOR (rs11121704) та ATG5 (rs510432) в розвитку алергічних захворювань у дітей
Авторы: Волосовець О.П., Досенко В.Є., Кривопустов С.П., Павлик О.В., Ємець О.В., Строй Д.О. — Національний медичний університет імені О.О. Богомольця, м. Київ
Рубрики: Педиатрия/Неонатология
Разделы: Клинические исследования
Версия для печати
Мета цього дослідження — визначення зв’язку однонуклеотидних поліморфізмів в генах mTOR (mammalian target of rapamycin; мішень рапаміцину у ссавців) і ATG5 (autophagy related gene 5; ген автофагії 5) та розвитку алергічного фенотипу у дітей. Генотипування mTOR (rs11121704) та ATG5 (rs510432) проводили в групі хворих з атопічними захворюваннями і контрольній групі за допомогою полімеразної ланцюгової реакції у реальному часі. Виявлено, що у 50 (54,9 %) пацієнтів і 43 (49,4 %) здорових дітей наявна мажорна алель (ТТ) поліморфізму rs11121704; 36 (39,6 %) і 32 (36,8 %) відповідно мали гетерозиготну алель (ТС); у 5 (5,5 %) і 12 (13,8 %) виявлено мінорну алель (СС). Варіанти з генотипом СС rs11121704 у гені mTOR визначалися у 2,5 раза частіше у контрольній групі, ніж серед хворих. Алельні варіанти в гені ATG5 розподілилися так: 51 (52 %) пацієнт та 43 (44,3 %) здорових дитини мали гетерозиготну алель (СТ), 29 (29,6 %) і 21 (21,6 %) відповідно мав мінорну алель (ТТ), у 18 (18,4 %) та 33 (34 %) була мажорна алель (СС). ТТ-генотип асоціюється з підвищеним ризиком ранньої маніфестації бронхіальної астми (до 3 років життя). Вважаємо, що поліморфізми в генах mTOR (rs11121704) та ATG5 (rs510432) можуть служити корисними прогностичними маркерами для оцінки ризику виникнення бронхіальної астми та іншої алергічної патології у дітей.
Цель данного исследования — определение связи однонуклеотидных полиморфизмов в генах mTOR (mammalian target of rapamycin; мишень рапамицина у самцов) и ATG5 (autophagy related gene 5; ген аутофагии 5) с развитием аллергического фенотипа у детей. Генотипирование mTOR (rs11121704) и ATG5 (rs510432) проводили в группе больных с атопическими заболеваниями и контрольной группе с помощью полимеразной цепной реакции в реальном времени. Обнаружено, что у 50 (54,9 %) пациентов и 43 (49,4 %) здоровых детей присутствовала мажорная аллель (ТТ) полиморфизма rs11121704, 36 (39,6 %) и 32 (36,8 %) соответственно имели гетерозиготную аллель (ТС), 5 (5,5 %) и 12 (13,8 %) — минорную аллель (СС). Варианты с генотипом СС rs11121704 в гене mTOR оказались в 2,5 раза чаще в контрольной группе, чем среди больных. Аллельные варианты в гене ATG5 распределились следующим образом: у 51 (52 %) пациента и 43 (44,3 %) здоровых детей присутствовала гетерозиготная аллель (СТ), 29 (29,6 %) и 21 (21,6 %) соответственно имели минорную аллель (ТТ), 18 (18,4 %) и 33 (34 %) — мажорную аллель (СС). ТТ-генотип ассоциируется с повышенным риском ранней манифестации бронхиальной астмы (до 3 лет жизни). Считаем, что полиморфизмы в генах mTOR (rs11121704) и ATG5 (rs510432) могут служить полезными прогностическими маркерами для оценки риска возникновения бронхиальной астмы и другой аллергической патологии у детей.
The objective of this study — to determine the association between single-nucleotide polymorphisms in mTOR (mammalian target of rapamycin) and ATG5 (autophagy related gene 5) genes and allergic phenotype in children. Genotyping for mTOR (rs11121704) and ATG5 (rs510432) was performed in group of patients with atopic diseases and in control group using real-time polymerase chain reaction. We have found that 50 (54.9 %) patients and 43 (49.4 %) healthy children had major allele (TT) of rs11121704 polymorphism; 36 (39.6 %) and 32 (36.8 %), respectively, had heterozygous allele (TC); 5 (5.5 %) and 12 (13.8 %) had minor allele (CC). Variants with CC genotype of rs11121704 in mTOR gene were revealed 2.5 times more frequently in the control group than among patients. Allelic variants in ATG5 gene were as follows: 51 (52 %) patients and 43 (44.3 %) healthy children had heterozygous allele (CT), 29 (29.6 %) and 21 (21.6 %), respectively, had minor allele (TT), and 18 (18.4 %) and 33 (34 %) had major allele (CC). TT genotype is associated with increased risk of early manifestation of bronchial asthma (until 3 years of life). We believe that polymorphisms in mTOR (rs11121704) and ATG5 (rs510432) genes may serve as important predictive markers in evaluating the risk of bronchial asthma and other allergic diseases in children.
однонуклеотидний поліморфізм, автофагія, mTOR, ATG5, алергічна патологія, діти.
однонуклеотидный полиморфизм, аутофагия, mTOR, ATG5, аллергическая патология, дети.
single nucleotide polymorphism, autophagy, mTOR, ATG5, allergic diseases, children.
Статья опубликована на с. 5-11
Вступ
Актуальність алергічної патології у дітей, а саме бронхіальної астми, атопічного дерматиту (АД) та інших атопічних захворювань зростає щороку. Кількість таких дітей в Україні становить 5–10 %, що можна порівняти з літературними даними щодо поширення цієї патології у світі. Так, у розвинених країнах рівень поширеності атопічних захворювань серед дитячого населення становить 15–30 % (Roider E. et al., 2013).
Зрозуміло, що ендогенним базисом розвитку алергічних захворювань є генетична схильність до порушення рівноваги в імунній системі, у взаємодії між різними типами лімфоцитів. Відомо, що диференціація ефекторних Т-клітин запускається під впливом зовнішніх антигенів і потребує одночасної взаємодії цитокінів, які забезпечують стимуляцію Т-клітинного рецептора (TCR) і, як наслідок, значних морфологічних і функціональних змін наївних Т-клітин [5, 17].
Існує принаймні шість різних ефекторних клонів з їх унікальними особливостями: Th1, Th2, Th9, Th17, Th22 і CD4+CD25+ регуляторні Т-клітини (Treg). Treg-клітини можуть походити з двох джерел. Природні Treg-клітини (nTreg) утворюються в процесі онтогенезу тимуса, в той час як індуковані Treg-клітини (iTreg) диференціюються з наївних периферичних Т-клітин при їх активації за наявності високої концентрації трансформуючого фактора росту бета (TGF-бета) і інтерлейкіна-2 (IL-2). Баланс між ефекторними Th- і Treg-клітинами може критично вплинути на результати багатьох захворювань людини. Підвищення рівня IL-6 і TGF-бета в мікросередовищі можуть спровокувати утворення Th17-клітин. Активація Т-клітин IFN-y і ІЛ-12 призводить до дозрівання Th1-клітин-ефекторів, присутність IL-4, IL-25, IL-33 і тимічного стромального лімфопоетину (TSLP) призводить до диференціації в Th2-клітини [1, 5, 17].
Функції різних типів клітин визначені цитокінами, які вони виробляють під час антигенної стимуляції. У патогенезі алергічного запалення секреція IL-4, IL-5, IL-31 і IL-13 Th2-клітинами призводить до алерген-специфічного синтезу імуноглобуліну Е (IgE), активації диференціювання еозинофілів та в результаті до дегрануляції тучних клітин [1].
Поряд із позаклітинними цитокіновими сигналами в імунній стимуляції лімфоцитів важливе значення мають внутрішньоклітинні механізми, зокрема автофагічного каскаду. Так, дослідниками (Delgofe et al.) була вивчена здатність сигнального шляху mTOR (mammalian target of rapamycin; мішень рапаміцину у ссавців) модулювати імунну відповідь та клітинну проліферацію. Мішень рапаміцину у ссавців (mTOR, також відомий як FRAP, RAFT, або RAPT) є еволюційно консервативною серин-треоніновою протеїнкіназою, що регулює клітинний метаболізм, ріст і проліферацію відповідно до зовнішніх чинників — доступність поживних речовин, вплив цитокінів [17]. З огляду на значну роль функцій mTOR у процесі клітинної диференціації було висловлено припущення, що mTOR у наївних Т-клітинах може визначати результат розпізнавання антигенів і Т-клітинну диференціацію.
Delgofe et al. досліджували роль у диференціації ефекторних Т-клітин і виявили, що mTOR-дефіцитні CD4 + Т-клітини не в змозі активувати специфічні транскрипційні фактори, такі як GATA-3, Т-bet, і RORgt. Крім того, автори показали, що нездатність до диференціювання в ефекторні клітини була обумовлена ослабленням відповідних STAT-сигналів у відповідь на поляризуючі цитокіни. Замість цього після активації mTOR-дефіцитні CD4+ Т-клітини диференціювалися в регуляторні клітини Foxp3+ [5]. У наступній роботі [6] показано, що Т-клітини, позбавлені mTORC1-активності, ще були в змозі стати Th2-клітинами. Автори досліджували вплив сигналізації mTORC2 на розвиток Th2. При видаленні сигналізації mTORC2 в Т-клітинах виявлено, що вони були не в змозі стати Th2-клітинами, але зберігалася їх здатність диференціюватися в Th1- і Th17-клітини. Ці дані визначають різні сигнальні шляхи як механізму, за допомогою якого mTOR регулює диференціювання Т-хелперів.
Становить інтерес те, що mTOR також бере участь у процесі росту, дозрівання тучних клітин і секреції цитокінів. Shin et al. у своєму дослідженні встановили, що туберозний склерозний комплекс 1 (TSC1) є критичним регулятором для сигналізації mTOR в тучних клітинах, підлеглих FcRI і С-Kit, і диференційно контролює дегрануляцію тучних клітин і продукцію цитокінів. TSC1-дефіцит призводить до порушення дегрануляції тучних клітин, але при цьому посилюється продукція цитокінів in vitro і в природних умовах після FcRI контакту. TSC1 має вирішальне значення для виживання тучних клітин через множинні шляхи апоптозу (в тому числі інгібуючої регуляції р53, miR-34a, активних форм кисню) і регуляцію Bcl-2 [20]. Крім того, сигналізація mTOR може модулювати ангіогенез і лімфангіогенез, що відіграють важливу роль у патогенезі астми. Інгібування mTOR блокує синтез фактора росту ендотелію судин (VEGF), фактор ангіогенезу, що індукує фенотип астми у мишей [8].
Як було зазначено, стимулювання mTORC1 знижує автофагію — процес внутрішньоклітинної деградації лізосомальних білків, цитоплазматичних компонентів, органел, що забезпечує механізм контролю якості білка. Відомо, що ініціація процесу автофагії відбувається під впливом щонайменше двох кіназ — ULK1 (відома також як ATG-1) та VPS34 (вакуольний протеїн 34). Як наслідок їх активації відповідні протеїни добудовуються в мембрану фагосоми, що завершує процес формування та дозрівання автофагосоми. mTORC1, при наявності відповідних умов, пригнічує активацію ULK1 шляхом фосфорилювання і таким чином стає негативним регулятором автофагії [7].
Автофагія відіграє важливу роль у функціонуванні імунної системи, а саме забезпечує знешкодження внутрішньоклітинних бактерій та вірусів, антигенпрезентацію на різних рівнях, проліферацію Т- і В-клітин, підтримку їх внутрішньоклітинного гомеостазу.
Відомо, що макроавтофагія після деградації протеїнів індукує транспортування їх до МНС (головного комплексу гістосумісності) ІІ типу в тимічних епітеліальних клітинах, що є необхідним для позитивної і негативної селекції лімфоцитів у тимусі [8, 20].
Аналогічно забезпечується презентація антигенів внутрішньоклітинних патогенів у комплексі МНС ІІ антигенпрезентуючих клітин СD4+-клітинам. Автофагія відповідає за процес антигенпрезентації у В-клітинах, макрофагах, дендритних клітинах. Антигени, що утворюють сильний зв’язок з Т-клітинним рецептором СD4 наївних Т-клітин, призводять до диференціювання їх у Th1- і Th17-клітини, тоді як низькоафінні комплекси «пептид-МНС/Т-клітинний рецептор» сприяють диференціації в Th2-клітини [4]. Таким чином, активність та функціонування Т-клітинного рецептора на поверхні Т-лімфоцитів після завершення процесів дозрівання та селекції, як і сам процес деградації протеїнів, можуть сприяти формуванню Th2-фенотипу. Важливо відзначити, що і цитокіни, притаманні Th1- та Th2-фенотипам, у свою чергу, регулюють активацію автофагії. Так, інтерферон y індукує протеоліз, а IL-4 та IL-13 пригнічують автофагію [15].
Окрім того, автофагія забезпечує деградацію мітохондрій та окислених протеїнів. Оксидативний стрес під час дизрегуляції протеолізу може призвести до ураження тканин організму. Наприклад, в разі бронхіальної астми рівні H2O2 та NO у повітрі, що видихається пацієнтом, прямо взаємопов’язані з тяжкістю перебігу астми [11].
Існує чіткий взаємозв’язок між процесами автофагії та активацією рецепторів розпізнавання патернів (PRRs), у тому числі Toll-, Nod- та RIG-I-подібних рецепторів. Так, активація рецепторів спричинює формування інфламасом, внаслідок чого завершується дозрівання IL-1b та IL-18 [14]. Крім того автофагія забезпечує дозрівання і вихід секреторних гранул тучних клітин, ремоделювання дихальних шляхів через збільшення виділення колагену з фібробластів [13].
Виходячи з наведених даних, ми припустили, що поліморфізми в генах mTOR та ATG5 можуть бути пов’язані з розвитком алергічного фенотипу в дитячому віці. Для перевірки цієї гіпотези ми досліджували відмінності в частоті SNPs (rs11121704 та rs510432) у дітей без алергічної патології (контрольна група) та дітей з бронхіальною астмою і/або іншою алергічною патологією, а також взаємозв’язок між цими двома поліморфізмами.
Матеріали та методи
Поліморфізм rs11121704 в гені mTOR досліджувався у 91 дитини, поліморфізм rs510432 в гені ATG5 — у 98 дітей віком від 5 до 18 років із бронхіальною астмою та/або іншим алергічним захворюванням, які перебували на стаціонарному лікуванні в алергологічному відділенні Київської міської дитячої клінічної лікарні № 2. Групу контролю становили 97 дітей віком від 5 до 18 років без алергічної патології для rs510432 в гені ATG5 та 87 дітей для rs11121704 в гені mTOR. Встановлення діагнозу АД відбувалося на підставі даних, внесених батьками в адаптовану затверджену анкету (ISAAC), і включало в себе скарги на висип, сухість шкіри, лущення, свербіж упродовж дитинства. Наявність бронхіальної астми встановлювалась на основі даних зі слів батьків щодо персистуючого свистячого дихання (≥ 2 епізодів нападів, не пов’язаних з інфекцією верхніх дихальних шляхів), даних спірометрії (знижений об’єм наприкінці першої секунди форсованого видиху (FEV1) і співвідношення FEV1/FVC (форсована життєва ємкість легень), позитивний тест з бета-2-агоністами), підвищення рівня IgE, позитивні шкірні прик-тести з аероалергенами (табл. 1).
/7/7.jpg)
Шкірні прик-тести були проведені та інтерпретовані згідно з протоколом Global Allergy and Asthma European Network, з використанням загальної панелі інгаляційних алергенів на підставі опублікованих методичних рекомендацій (документ Європейської академії алергології та клінічної імунології (EAACI) та протокол «Північні стандарти і міжнародне дослідження астми та алергії серед дітей» (ISAAC) ІІ фаза). Батьки підписали інформовану згоду до включення в дослідження. Окрім симптомів астми, АД на момент огляду або в анамнезі, алергічного риніту, в усіх дітей був принаймні один позитивний результат шкірного прик-тесту з панелі з 15 алергенами.
Вибір SNP (однонуклеотидного поліморфізму)
Поліморфізми rs11121704 у гені mTOR та rs510432 у гені ATG5 були обрані для генотипування, тому що гіпотетично вони можуть впливати на розвиток алергічного фенотипу і, як повідомляється, зустрічаються в європейських популяціях.
Виділення дезоксирибонуклеїнової кислоти (ДНК)
Забір букального епітелію проводився з використанням букальних щіток із подальшим заморожуванням зразків та їх зберіганням при температурі –20° С. ДНК для генотипування екстрагували зі зразків, використовуючи набори DiatomTM Prep 200 («Лаборатория Изоген», РФ) відповідно до протоколу виробника. Концентрацію ДНК визначали за допомогою спектрофотометра NanoDrop ND1000 (NanoDrop Technologies Inc., США).
Полімеразна ланцюгова реакція
Реакції ампліфікації проводили з використанням Fast Real-time PCR System (Applied Biosystems, США), в кінцевій реакції об’ємом 20 мкл, що містив 2X TaqMan Універсальний Master Mix (Applied Biosystems, США), assay C_910351_10 для поліморфізму rs510432 в гені ATG5 і матричну ДНК. Ампліфікація фрагментів генів складалася зі стадії денатурації при 95° С протягом 20 с, а потім 40 циклів ампліфікації при 95° С протягом 3 с і 60° С протягом 30 с. Аналіз даних проводився з 7500 Fast Real-Time PCR Software.
Статистичний аналіз
Отримані дані обробляли статистично з використанням програми Excel 2000 та Оrigin 7.0. Відмінність в частоті генотипу і алелей між групою пацієнтів із бронхіальною астмою і контрольною групою визначалась за допомогою χ2-тесту Пірсона. Р-значення менше 0,05 (р < 0,05) вважалося статистично значущим. Для визначення та підтвердження взаємодії між SNP, визначення типу цієї взаємодії використовувався метод багатофакторного зменшення розмірності (MDR — Multofactorial Dimentionality Reduction), а також графічне зображення епістазу за допомогою побудови дендрограм взаємодії. Для перевірки рівноваги Харді — Вайнберга використовували SNPAnalyzer (веб-програмне забезпечення).
Результати
У 72 (73,4 %) дітей було загострення бронхіальної астми, що проявлялось експіраторною задишкою, свистячими хрипами і кашлем. У 40 (55,5 %) пацієнтів діагностовано легкий ступінь загострення, 31 (43,1 %) мали загострення середньої тяжкості, 1 (1,4 %) — тяжке загострення, 28 % дітей з діагнозом бронхіальної астми в стадії ремісії проходили планове обстеження. Загалом у 60 (83,3 %) пацієнтів виявлено зниження показника FEV1 (< 80 %), 34 (47,2 %) мали позитивний тест з бета-2-агоністом. У сімейному анамнезі всіх дітей були випадки атопічних захворювань. 98 (100 %) дітей мали АД в анамнезі, з них 35 (35,7 %) дітей упродовж останнього часу відмічають прояви АД у вигляді висипу, сухості шкіри, лущення, свербежу, ліхенізаціі. У 34 (34,7 %) дітей прояви АД відзначались із дня народження до 1 року, у 16 (16,3 %) — до 3 років, у 8 (8,2 %) — до 7 років, у 7 (7,2 %) — до 10 років, у 5 (5,1 %) прояви з’явились після року.
Інша супутня алергологічна патологія визначалась у 46 (46,9 %) дітей. У 23 (23,5 %) дітей мав місце алергічний риніт: із них 18 (18,4 %) мали цілорічний алергічний риніт, 5 (5,1 %) — сезонний. Серед алергологічних захворювань в анамнезі відзначалась гостра алергічна кропив’янка — 4 (4,1 %) дітей, харчова алергія — 9 (9,2 %) дітей.
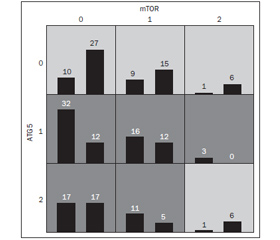
Генотипування показало такий розподіл алельних варіантів у гені mTOR (рис. 1).
/8/8.jpg)
Розподіл генотипів rs11121704 у гені mTOR відповідає закону Харді — Вайнберга. Виявлено, що у 50 (54,9 %) пацієнтів і 43 (49,4 %) здорових дітей була наявна мажорна алель (ТТ) (р > 0,05 за χ2-тестом); 36 (39,6 %) і 32 (36,8 %) пацієнтів та здорових дітей відповідно мали гетерозиготну алель (ТС) (р > 0,05 за χ2-тестом, відношення шансів (ОR) 0,97 (95% довірчий інтервал (ДІ) 0,52–1,81) та 5 (5,5 %) і 12 (13,8 %) мали мінорну алель (СС) (р > 0,05 за χ2-тестом, ОR 0,36 (95% ДІ 0,11–1,05) (табл. 2–4). Варіанти з генотипом СС rs11121704 у гені mTOR виявилися в 2,5 раза частіше у контрольній групі, ніж серед хворих.
/8/8_2.jpg)
Розподіл алельних варіантів у гені ATG5 наведено на рис. 2.
/9/9.jpg)
Розподіл генотипів rs510432 в гені ATG5 також відповідає закону Харді — Вайнберга. Виявлено, що у 51 (52 %) пацієнтів та 43 (44,3 %) здорових дітей була гетерозиготна алель (СТ) (р < 0,05 за χ2-тестом, ОR 2,53 (95% ДІ 1,14–5,74) у 29 (29,6 %) пацієнтів та у 21 (21,6 %) дітей із контрольної групи наявна мінорна алель (ТТ) (р < 0,05 за χ2-тестом, ОR 1,00) та 18 (18,4 %) пацієнтів і 33 (34 %) здорових дітей мали мажорну алель (СС) (р < 0,05 за χ2-тестом, ОR 2,17 (95% ДІ 1,09–4,46) (табл. 2–4). ТТ-генотип поліморфізму rs510432 в гені ATG5 асоціюється з підвищеним ризиком ранньої маніфестації бронхіальної астми (до 3 років життя) (р = 0,005369) (табл. 5).
Серед дітей із поліморфізмом rs11121704 в гені mTOR було виявлено, що у 30 (32,9 %) хлопчиків і 20 (21,9 %) дівчат була мажорна алель, 23 (25,3 %) і 13 (14,3 %) хлопчиків та дівчат, відповідно, мали гетерозиготну алель, 1 (1,1 %) і 4 (4,4 %) — мінорну алель (СС) (р > 0,05 за χ2-тестом). Розподіл за статтю у дітей із поліморфізмом в гені ATG5 відбувся таким чином: 29 (29,6 %) і 22 (22,5 %) хлопчиків та дівчат, відповідно, мали гетерозиготну алель; 13 (13,3 %) і 13 (13,3 %) — мінорну алель (СС); у 14 (14,3 %) хлопчиків і 7 (7,1 %) дівчат була мажорна алель (р > 0,05 за χ2-тестом).
Чітко видно (рис. 3), що між поліморфізмами ATG5 та mTOR не спостерігається ефекту взаємодії і вони представляють незалежні головні ефекти.
/10/10.jpg)
Обговорення та висновки
Літературні дані свідчать про значну роль mTOR у регуляції сигналів цитокінів, диференціюванні й розвитку ефекторних Th-клітин, дозріванні й дегрануляції тучних клітин, регуляції процесу презентації антигену шляхом автофагії, що згодом призводить до формування Th2-клітинного фенотипу. Сигнальний шлях mTOR також є важливим у функціонуванні нейтрофілів, моноцитів, дендритних клітин, В- та Т-клітин. Попередні дослідження визначили mTOR як критичний регулятор клітинного метаболізму, даючи початок розвитку нових генетичних моделей, щоб показати важливу роль цього шляху в гомеостазі та функції Т-лімфоцитів.
Ідентифікація факторів ризику, що можуть вплинути на ризик розвитку атопічної патології, дуже важлива. Останнім часом ряд «випадок — контроль» досліджень показав зв’язок поліморфізмів в гені mTOR з ризиком розвитку раку [2, 3, 9, 10, 12, 19], зокрема, поліморфізму rs11121704 (T > C), широко вивченого в популяціях Європи, США та Китаю. Дослідниками був показаний зв’язок TT-генотипу rs11121704 з клінічними параметрами (OR = 1,53, 95% ДІ 1,01–2,32, р = 0,044), такими, як ризик смертності, метастазів і стійкість до хіміотерапії [13]. Також було показано, що поліморфізм rs2295080 в промоторі зменшує транскрипційну активність у дослідженні in vitro і пов’язаний з низькою експресією мРНК mTOR у нирковій тканині [12]. В описаному дослідженні функціональне значення поліморфізму rs11121704 було визначене за допомогою SNPexp онлайн-інструменту для оцінки можливого біологічного впливу на експрессію гена mTOR. Випадки з ТТ-генотипом мали вищі рівні експресії гена mTOR, ніж ТС- і СС-генотипи, хоча і не було досягнуто рівня статистичної значущості (р = 0,059).
Поліморфізми rs11121704 в гені mTOR та rs510432 в гені ATG5 були широко вивчені в європейській популяції і, отже, були обрані нами для дослідження. Розподіл варіантів поліморфізму rs11121704 в гені mTOR в європейській популяції: генотип ТТ — 52,7 %, генотип ТС — 41,1 %, генотип СС — 6,2 %, алель Т — 73 %, алель С — 27 %. Розподіл варіантів поліморфізму гена ATG5 rs510432 в європейській популяції: генотип СС — 31,9 %, генотип СТ — 44,2 %, генотип ТТ — 23,9 %, алель С — 54 %, алель Т — 46 %.
Ми досліджували взаємозв’язок дитячих атопічних захворювань і mTOR регуляції та автофагії. Було виявлено можливий зв’язок між SNP у гені mTOR та ATG5 і алергічним фенотипом у дітей. Варіанти з генотипом СС rs11121704 в гені mTOR зустрічалися у 2,5 раза частіше у контрольній групі, ніж серед хворих. Також було знайдено статистично значущі відмінності у розподілі генотипів rs510432 в гені ATG5 у контрольній групі та групі пацієнтів. У дітей з алергічною патологією варіанти з ТТ-генотипом зустрічаються частіше, ніж у групі контролю.
Асоціації генотипів із клінічними параметрами алергічних захворювань не було встановлено в попередніх дослідженнях. У цьому дослідженні нами було виявлено, що ТТ-генотип поліморфізму rs510432 в гені ATG5 асоціюється з підвищеним ризиком ранньої маніфестації бронхіальної астми (до 3 років життя) (р = 0,005369) (табл. 5).
Між поліморфізмами ATG5 та mTOR не спостерігається ефекту взаємодії і вони представляють незалежні головні ефекти (рис. 3).
Зважаючи на ключову роль лізосомного протеолізу в реалізації та регуляції імунної відповіді процес автофагії в патогенезі алергічних захворювань достатньо широко вивчений. Останні дослідження продемонстрували взаємозв’язок між поліморфізмами гена ATG5 (rs 12201458 та rs 510432) та бронхіальною астмою у дітей в американській популяції. Мінорна алель (Т) гена ATG5 rs 510432 пов’язана з підвищеним ризиком бронхіальної астми у дітей. Окрім того, як показано в експерименті, поліморфізм rs510432 підвищує активність промотора гена: варіант алелі С призводив до підвищення активності промотора на 23 % порівняно з варіантом алелі Т. На наступній стадії дослідження було підтверджено прогнозоване підвищення експресії гена ATG5 у пацієнтів із бронхіальною астмою у стадії загострення [16]. Також було виявлено залежність показників спірограми (FEV1) від поліморфізму гена ATG5 (rs 12212740) у пацієнтів із бронхіальною астмою в американській та канадській популяціях [18].
Протеїн ATG5, що є одним із основних структурних білків автофагосоми, часто обирається як об’єкт дослідження різноманітних захворювань. Так, підвищений рівень експресії гена ATG5 спостерігається при хворобі Паркінсона, виразковій хворобі шлунка. Інгібування автофагії являє собою потенційно новий метод лікування нейродегенеративних, онкологічних, метаболічних, інфекційних та атопічних захворювань.
Таким чином, поліморфізми генів mTOR та ATG5 можуть розглядатися як перспективний генетичний маркер для оцінки ризику виникнення бронхіальної астми та іншої алергічної патології у дітей.
1. Brand E.B., Sivaprasad U. Th2 Cytokines and Atopic Dermatitis // J. Clin. Cell. Immunol. — 2011. — 2(3). — 110.
2. Cao Q., Ju X., Li P., Meng X., Shao P. et al. A functional variant in the MTOR promoter modulates its expression and is associated with renal cell cancer risk // PLoS One. — 2012. — 7(11). — e50302. doi: 10.1371/journal.pone.0050302.
3. Chen J., Shao P., Cao Q., Li P., Li J. et al. Genetic variations in a PTEN/AKT/mTOR axis and prostate cancer risk in a Chinese population // PLoS One. — 2012. — 7(7). — e40817. doi: 10.1371/journal.pone.0040817.
4. Datta S., Milner J.D. Rostrum. Altered T-cell receptor signaling in the pathogenesis of allergic disease // J. Allergy Clin. Immun. — 2011 — 127(2). — 351-354. Doi:10.1016/j.jaci.2010.11.0339.
5. Delgoffe G.M., Kole T.P., Zheng, Y., Zarek P.E., Matthews K.L., Xiao B., Worley P.F., Kozma S.C., Powell J.D. mTOR differentially regulates effector and regulatory T cell lineage commitment // Immunity. — 2009. — 30(6). — 832-844. doi: 10.1016/j.immuni.2009.04.014.
6. Delgoffe G.M., Pollizzi K.M., Waickman A.D., Heikamp E., Meyers D.J., Horton M.R., Bo Xiao, Worley P.F., Powell J.D. The mammalian Target of Rapamycin (mTOR) regulates T helper cell differentiation through the selective activation of mTORC1 and mTORC2 signaling // Nature immunology. — 2011. — 12(4). — 295-303.
7. Dunlop E.A., Tee A.R. mTOR and autophagy: A dynamic relationship governed by nutrients and energy // Semin. Cell. Dev. Biol. — 2014. http://dx.doi.org/10.1016/j.semcdb.2014.08.006
8. Fredriksson K., Fielhaber J.A., Lam J.K., Yao X., Meyer K.S. et al. Paradoxical Effects of Rapamycin on Experimental House Dust Mite-Induced Asthma // PLoS One. — 2012. — e33984. doi:10.1371/journal.pone.0033984.
9. He J., Wang M.Y., Qiu L.X., Zhu M.L., Shi T.Y. et al. Genetic variations of mTORC1 genes and risk of gastric cancer in an eastern chinese population // Mol. Carcinog. — 2013. — 52 Suppl. — 170-179. doi: 10.1002/mc.22013.
10. Hildebrandt M.A., Yang H., Hung M.C., Izzo J.G., Huang M. et al. Genetic variations in the PI3K/PTEN/AKT/mTOR pathway are associated with clinical outcomes in esophageal cancer patients treated with chemoradiotherapy // J. Clin. Oncol. 2009. — 27. — 857-871. doi: 10.1200/jco.2008.17.6297.
11. Horvath I., Donnelly L.E., Kiss A. Combined use of exhaled hydrogen peroxide and nitric oxide in monitoring asthma // Am. J. Respir. Crit. Care Med. — 1998. — 158. — 1042-1046.
12. Huang L., Huang J., Wu P., Li Q., Rong L. et al. Association of genetic variations in mTOR with risk of childhood acute lymphoblastic leukemia in a Chinese population // Leuk. Lymphoma. — 2012. — 53. — 947-951. doi: 10.3109/10428194.2011.628062.
13. Jianbo Shao, Ying Li, Peiwei Zhao, Xin Yue, Xiaohui Liang Xuelian He. Association of mTOR Polymorphisms with Cancer Risk and Clinical Outcomes: A Meta-Analysis // PLoS One. — 2014 May 9. — 9(5). — e97085. DOI: 10.1371/journal.pone.0097085.
14. Jun A., Hiromichi H., Kazuyoshi K. Autophagy in the Pathogenesis of pulmonary disease // Intern. Med. — 2013. — 52. — 2295-2303.
15. Jyothula S.S., Eissa N.T. Autophagy and role in asthma // Curr. Opin. Pulm. Med. — 2013. — 19. — 30-35.
16. Martin L.J., Gupta J., Jyothula S.S. Functional variant in the autophagy-related gene 5 gene promoter is associated with childhood asthma // PLoS One. — 2012. — 7. — e33454.
17. O’Brien T.F., Xiao-Ping Zhong. The Role and Regulation of mTOR in T Lymphocyte Function // Arch. Immunol. Ther. Exp (Warsz). — 2012 Jun. — 60(3). — 173-181.
18. Poon A.H., Chouiali F., Tse S.M. Genetic and histologic evidence for autophagy in asthma pathogenesis // J. Allergy Clin. Immun. — 2012. — 129. — 569-571.
19. Pu X., Hildebrandt M.A., Lu C., Lin J., Stewart D.J. et al. PI3K/PTEN/AKT/mTOR pathway genetic variation predicts toxicity and distant progression in lung cancer patients receiving platinum-based chemotherapy // Lung. Cancer. — 2011. — 71. — 82-88. doi: 10.1016/j.lungcan.2010.04.008.
20. Shin J., Pan H. Xiao-Ping Zhong Regulation of mast cell survival and function by tuberous sclerosis complex 1 // Blood. — 2012 Apr 5. — 119(14). — 3306-3314.